
Fig. 1 (from ref. 5). Outlines of the statolith complexes in central cress statocytes (stories II and III) from ground control (gc), flight control (fc) and flight probe (fp).

Two experiments with cress seedlings were performed to investigate the position of statoliths under microgravity conditions. For this purpose, cress seedlings were grown for 26 hr under 1 g conditions and then subjected to 'µg for 6 min. For documentation of the statoliths' position, seedlings were chemically fixed at the beginning (flight control) and the end (flight probe) of the 'µg phase. Vertically grown seedlings from the laboratory were used as additional controls (ground control). The distance of statoliths from the originally lower cell wall was estimated using semi-thin sections. Both controls did not show any significant difference, indicating no notable influence by launch conditions on the statoliths' position. On the other hand, there was a significant difference between the flight control and flight probe, revealing a displacement of statoliths of approximately 3 'µm in the direction opposite to the originally acting gravity vector. One can conclude from these results that the position of statoliths depends on the gravitational field and on intracellular forces probably exerted by elements of the cytoskeleton.
The statolith theory of Nemec and Haberlandt predicts that intracellular dense particles such as starch-filled plastids trigger the gravitropic stimulus response chain. After deviation from the vertical, these particles show displacement depending on the gravity vector (see ref. 1). After incubation of roots with the fungal toxin cytochalasin, an inhibitor of actin filaments, sedimentation of statoliths is more pronounced in comparison to untreated controls.² Affinity labelling by rhodamin phalloidin of the statocytes from coleoptiles³ and hypocotyls4 revealed a network of actin filaments surrounding the statoliths. These results suggest that the position of statoliths depends on two forces: the force exerted by the gravitational field and tensional forces caused by cytoskeletal elements, especially actin filaments. In order to test this working hypothesis, experiments were performed during the parabolic flights of the TEXUS 18 & 19 rockets.
Seedlings of cress (Lepidium sativum L.) were cultivated for 20 hr in the normal vertical position in darkness at 25°C. For reasons of adaptation, seedlings of uniform length were transferred for 5 hr to seed holders, which were finally mounted in flight containers (Kayser-Threde, Munich, Germany) 1 hr before launch. For optical and electron microscopy, the seedlings were chemically fixed5 at the beginning and end of the 'µg phase by automatically flooding the growth containers. See ref. 6 for a detailed description of the flight hardware and the inflight data.
Cress seedlings growing for 26 hr under 1 g conditions were chemically fixed at the beginning (flight control) and the end (flight probe) of the 'µg phase of approximately 6 min. In addition, a second control was performed on Earth (ground control). Looking at the outlines of statolith complexes (Fig. 1), clear differences became evident between the ground control (gc), flight control (fc) and flight probe (fp).
Fig. 1 (from ref. 5). Outlines of the statolith complexes in
central cress statocytes (stories II and III) from ground control
(gc), flight control (fc) and flight probe (fp).
Minor differences between gc and fc were characterised by a more undulating outline of fc, indicating some disturbances during the launch phase. No difference was visible between gc and fc in the distance of statolith complexes from the original lower cell wall. With fp, however, this distance was remarkably larger, indicating displacement of the statolith complex in the opposite direction to the originally acting gravity vector.
Looking at individual statoliths of different stories in the root cap by optical microscopy (Fig. 2), it was possible to quantify the displacement of statoliths (Fig. 3). There was a significant displacement of statoliths in the range of 3-4 'µm during the short, 6 min, period of 'µg. This means that statoliths moved over a distance corresponding approximately to their diameter. On the basis of these results, a new model was suggested for transformation of the gravity stimulus by participation of cytoskeletal elements, especially actin filaments.7, 5

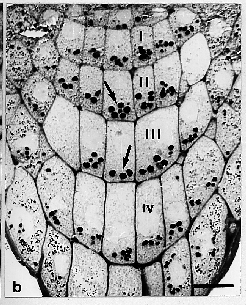

Fig. 2 (from ref. 5). Optical microscope photographs of median
longitudinal sections through the statenchyma of cress roots. a:
ground control (gc); b: flight control (fc); c: flight probe
(fp). Arrows indicate statoliths. Bars 20 µm.

Fig. 3 (from ref. 5). Mean distance of statoliths from the
originally lower cell wall in stories II and III of cress root
caps. Ground control (gc), flight control (fc), flight probe
(fp).
The authors wish to express their thanks to personnel from DLR and DASA for management and performance of the experiments, to personnel of Kayser-Threde for development of the hardware and support during the experiment procedure, to Petra Peters, Leo Schwarz, Horst Laubach and Paul Blaszcyk from the Botany Institute in Bonn for technical assistance and support with the hardware. Financial support to AGRAVIS (Bonn) by DARA (Bonn) and MWF (Düsseldorf) is gratefully acknowledged.



 SP1206
SP1206