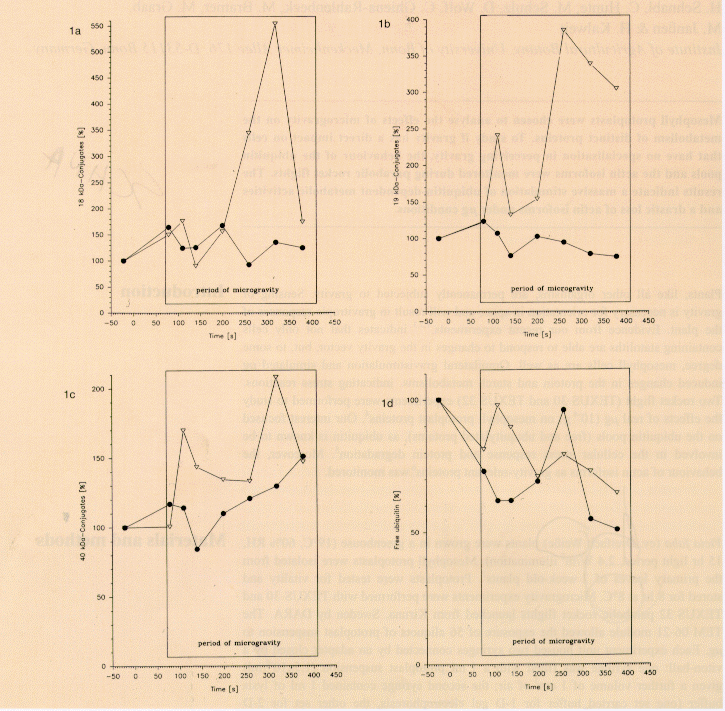
Figs. 1a-1d. Relative changes (%) of 18, 19 & 40 kDa ubiquitin-conjugates and free ubiquitin in mesophyll cell protoplasts from Vicia faba L. during the TEXUS 32 flight. Flight samples = triangles; ground control = circles; box = µg period.

Mesophyll protoplasts were chosen to analyse the effects of microgravity on the metabolism of distinct proteins. To study if gravity has a direct impact on cells that have no specialisation in perceiving gravity, the behaviour of the ubiquitin pools and the actin isoforms were monitored during parabolic rocket flights. The results indicate a massive stimulation of ubiquitin-dependent metabolic activities and a drastic loss of actin isoforms under µg conditions.
Plants, like all other organisms, are permanently subjected to gravity. Sensing of gravity is necessary for orientation in space and will result in gravitropic responses of the plant. Evidence from our recent experiments,1,2,3 indicates that not only cells containing statoliths are able to respond to changes in the gravity vector, but, to some degree, mesophyll cells are as well. Omnilateral gravistimulation and simulated µg induced changes in the protein and starch metabolisms, indicating stress reactions. Two rocket flight (TEXUS 30 and TEXUS 32) experiments were performed to study the effects of real µg (10-4g) on mesophyll protoplast proteins 4. Our interest focused on the ubiquitin pools (free and ubiquitylated proteins), as ubiquitin is known to be involved in the cellular stress response and protein degradation5. Moreover, the behaviour of actin isoforms as gravity-relevant proteins6 was monitored.
Vicia faba (cv. Dreifach Weiße) plants were grown
in a greenhouse (19°C, 60% RH, 15 hr light period, 2.4
W/m² illumination). Mesophyll protoplasts were isolated from
the primary leaves of 2 week-old plants7. Protoplasts
were tested for vitality and stored for 8 hr at 8°C.
Microgravity experiments were performed with TEXUS 30 and TEXUS
32 parabolic rocket flights launched from Kiruna, Sweden by DARA.
The TEM 06-21 module allowed the exposure of 56 aliquots of
protoplast suspension to µg. Each experiment unit housed two
syringes connected by an adapter closed by a (*)VITON®-ball: one
syringe was filled with a 1 ml protoplast suspension
(1.5x10-6/ml) given a further volume of 1 cm³ of
air; the second syringe contained 1 ml of lysis buffer (one set
carried buffer for 1-D gel electrophoresis, the other set for 2-D
electrophoresis).
(*) (VITON® is a registered trademark of Dupont Dow Elastomers)
Two hours before launch, 56 of these pre-cooled (10°C) units were mounted in the module in pre-cooled metallic holders. The temperature was slowly raised to 23°C before liftoff and maintained by Peltier elements. Protoplast samples were lysed during the flights and proteins extracted at set times. This was achieved by hydraulic pressure on the plug of the lysis buffer syringe. For TEXUS 30, one of each triplicated unit was lysed before launch (-22 s), shortly after the boost phase (78 s) and at the end of the µg phase (359 s). A larger module on TEXUS 32 allowed additional measurement points (109, 139, 199, 259, 319 s). 45 min after payload landing, the samples were recovered from the syringes, boiled for 10 min, centrifuged (10 000 g) and stored at -80°C. Proteins of flown and ground samples were separated by 1-D or 2-D electrophoresis and western-immunoblotted with polyclonal ubiquitin or monoclonal actin antibodies.
Densitometric quantification of ubiquitin-immunoblots of the separated protein extracts revealed changes within the ubiquitin pools (Figs. 1a-1d). An 18 kD ubiquitylated protein dramatically increased after about 125 s of µg, with the highest concentration at the 315 s measurement point before declining to a slightly increased level compared to pre-launch (-25 s) and launch (75 s) controls. The amount of this ubiquitinylated protein in the ground control samples oscillated during the examination time between 100% and 150%. A similar effect was observed quantifying a 19 kD conjugate. While ground control samples showed only a slight decrease, flight conditions resulted in a massive increase of this polypeptide, with peaks at 170 s and 300 s, the latter reaching 340%. A 40 kD ubiquitinylated protein exhibited similar behaviour. The oscillation of free ubiquitin indicates an activation of the ubiquitin system, where free ubiquitin is conjugated to target proteins and bond ubiquitin is finally released again by the activity of the 26S proteasom complex, which hydrolyses the target protein to amino acids. Accelerated ubiquitinylation of proteins, however, appears almost immediately in stressed organisms. The 18, 19 & 40 kD proteins that oscillate with remarkably higher maxima in the flight samples may present substrates for reversible ubiquitylation, such as ubiquitin-activating and ubiquitin- carrying enzymes. Although the identity of the proteins is still in question, they present a strong indication for highly activated ubiquitin-dependent activities in the protoplasts under µg conditions. Actin-based microfilaments are thought to be a factor in gravireception5. Vicia faba has four actin isoforms, which are constitutive polypeptides of the different tissues and organs8, and which show a distinct distribution within the specific cellular compartments: two isoforms (pI 5.9 and 6.0) were found in plasma membrane and tonoplast preparations, whereas the pI 5.95 and 6.05 polypeptides were present in the soluble fraction. Immunoblot analysis of actin isoforms during the parabolic rocket flights revealed a slight increase of the four major isoforms after the boost phase and a decrease during the µg phase. At the end of the µg phase, only the 6.0 and the 5.95 spots were still visible. Ground controls showed no changes (Figs. 2a-2d). The physiological meaning of this rapid decrease in actin isoforms remains unclear. One can speculate that a part of the isoforms becomes functionless under µg and undergoes degradation. More likely, actin isoform decline is a result of stress-induced proteolysis. It has been shown that the cellular organisation is also disturbed in gravity-insensitive cells. Those disturbances are believed to cause stress reactions influencing the protein metabolism as reflected by the µg-affected ubiquitin pools. Our results provide strong evidence that µg has a direct influence on mesophyll protoplast protein metabolism.
Figs. 1a-1d. Relative changes (%) of 18, 19 & 40 kDa
ubiquitin-conjugates and free ubiquitin in mesophyll cell
protoplasts from Vicia faba L. during the TEXUS 32 flight. Flight
samples = triangles; ground control = circles; box = µg
period.



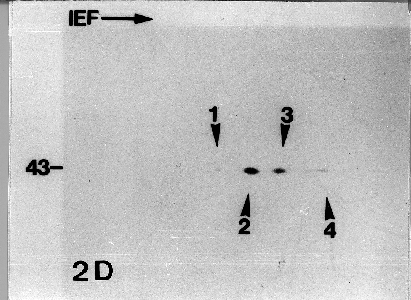
Figs. 2a-2d: Actin immunoblots of mesophyll cell protoplasts
from Vicia faba L. after 2-D SDS-PAGES. The molecular mass of the
immunostained spots is 43 kDa. Arrow heads mark the different
actin isoforms with pI values of (1) 6.05, (2) 6.0, (3) 5.95 and
(4) 5.9. 2a is before launch; 2b after the boost phase; 2c during
the µg phase; 2d at the end of the µg phase.
Financial support (AGRAVIS) by DARA (Bonn) and the Ministry of Science and Research (Düsseldorf, Nordrhein-Westfalen) to H. Schnabl is gratefully acknowledged.


 SP1206
SP1206